Agrees with our experimental observation that BRE-silencing accelerated chondrogenic differentiation of HUCPV cells. Because actin and ANXA2 play a role in cell migration, we investigated the effect of BRE-silencing on cell movement. The HUCPV cells showed reduced ability to migrate after BREsilencing. The reduction is probably associated with the downregulation of actin protein which affected the cytoskeleton to change dynamically during movement �C since migration requires a synchronized interactions of actin and its interacting partners such as actin-binding proteins and integrins. ANXA2 has also been implicated in the regulation of HSC binding to osteoblasts and homing to the bone marrow niche. Further studies will be required to establish whether BRE has a role in cell homing. BRE-silencing could also suppress CALR, FUBP1 and PKM2 expression. In mESCs, down-regulation of CALR resulted in reduction of calcium ion level to enhance adipogenic differentiation. It has also been reported that CALR may be involved in the regulation of osteogenesis and chondrogenesis in MSCs. CALR expression is reduced in the early stage of osteoblast differentiation in MC-3T3-E1 cells. Overexpression of CALR inhibits both the basal and vitamin D-induced expression of osteocalcin and calcium ion accumulation in the extracellular matrix and mineralization of bone nodules in cultures. These findings may 4-(Benzyloxy)phenol explain our observation of why BRE-silencing resulted in enhanced osteogenic and chondrogenic differentiation in HUCPV cells. BRE-silencing also negatively regulates FUBP1 and PKM2. FUBP1 is present in undifferentiated cells but not in differentiated cells and  regulates c-myc expression by binding to a single-stranded far-upstream element upstream of the c-myc promoter. Previous report showed that down-regulation of FUBP1 and subsequent down-regulation of c-myc were needed for lung cell differentiation in mice. This protein may act both as an activator and repressor of transcription. Likewise, PKM2 has been determined to stimulate Oct4-mediated transcriptional activation. Oct4 and c-myc constitute are two of the Yamanaka transcription factors crucial for maintaining pluripotency of stem cells. BRE-silencing resulted in decreased expression of pluripotency factors and may explain our observation of enhanced osteogenic and chondrogenic differentiation in HUCPV cells. Further studies are required to examine the effect of BRE-silencing on other mesenchymal Ginsenoside-F4 lineage differentiations. In sum, our findings suggest a multifunctional role for BRE in maintaining stemness and the cytoskeletal architecture of HUCPV cells. In addition, BRE expression can be manipulated to accelerate induced chondrogenic and osteogenic differentiation in HUCPV cells. Breast cancer is the most common malignant neoplasm and the second leading cause of death from cancer in women both in the USA and Europe. It is established that both adjuvant chemotherapy and hormonal therapy prolong disease-free and overall survival. Anthracyclines and taxanes are the two most commonly used classes of agents in this setting. However, despite the optimal management of patients with early-stage breast cancer, eventually approximately 30% of them suffer from disease relapse. Metastatic breast cancer is an incurable disease with few therapeutic options. With the increasing use of anthracyclines and taxanes in the adjuvant setting the number of available drugs for these patients is even more limited. Obviously, there is an unmet need for the introduction to the clinic of agents with novel modes of action, lack of cross-resistance with existing agents and promising activity in metastatic breast cancer.
regulates c-myc expression by binding to a single-stranded far-upstream element upstream of the c-myc promoter. Previous report showed that down-regulation of FUBP1 and subsequent down-regulation of c-myc were needed for lung cell differentiation in mice. This protein may act both as an activator and repressor of transcription. Likewise, PKM2 has been determined to stimulate Oct4-mediated transcriptional activation. Oct4 and c-myc constitute are two of the Yamanaka transcription factors crucial for maintaining pluripotency of stem cells. BRE-silencing resulted in decreased expression of pluripotency factors and may explain our observation of enhanced osteogenic and chondrogenic differentiation in HUCPV cells. Further studies are required to examine the effect of BRE-silencing on other mesenchymal Ginsenoside-F4 lineage differentiations. In sum, our findings suggest a multifunctional role for BRE in maintaining stemness and the cytoskeletal architecture of HUCPV cells. In addition, BRE expression can be manipulated to accelerate induced chondrogenic and osteogenic differentiation in HUCPV cells. Breast cancer is the most common malignant neoplasm and the second leading cause of death from cancer in women both in the USA and Europe. It is established that both adjuvant chemotherapy and hormonal therapy prolong disease-free and overall survival. Anthracyclines and taxanes are the two most commonly used classes of agents in this setting. However, despite the optimal management of patients with early-stage breast cancer, eventually approximately 30% of them suffer from disease relapse. Metastatic breast cancer is an incurable disease with few therapeutic options. With the increasing use of anthracyclines and taxanes in the adjuvant setting the number of available drugs for these patients is even more limited. Obviously, there is an unmet need for the introduction to the clinic of agents with novel modes of action, lack of cross-resistance with existing agents and promising activity in metastatic breast cancer.
Monthly Archives: May 2019
We demonstrated that the dimerization between the pathogenic variants also occurred with lower
Subsequently, the PCR fragments were purified and subcloned into the SalI and BamHI sites of the expression vector pEGFP-N1. The mammalian expression plasmids pTL-Flag-TPI-WT and pTL-Flag-TPI-Glu104Asp were generated by subcloning the corresponding DNA fragments from pACT-TPI-WT or pACTTPI-Glu104Asp into the XhoI/NotI sites of the vector pTL-FlagC, respectively. 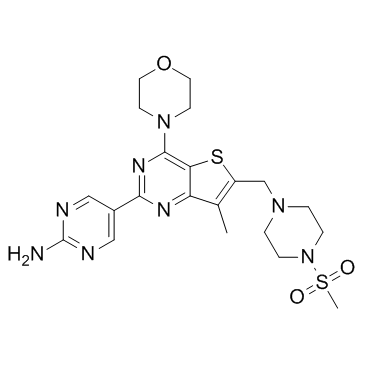 PCR was performed under conditions described earlier and all resultant products were verified by sequencing. As aforementioned, the frequency of TPI null alleles is much higher as the rare incidence of TPI deficiency, and in vitro measurements of a mutant TPI variant purified from E. coli demonstrated that the purified TPI protein carrying the Phe240Leu mutation exhibited a 6-fold higher activity than expected from the measurements performed with the patient erythrocyte extract. Moreover, theoretical calculations demonstrated that the extremely high DHAP level observed in patients’erythrocytes does not correlate with the measured residual TPI activity. Since patients’metabolites highly depend on the genetic background and environmental factors, we have generated an in vivo system Folinic acid calcium salt pentahydrate allowing analysis of the enzymatic activity of wild-type or pathogenic TPI variants inside a cell without additional side effects. At least three different isoelectric TPI variants encoded by the single TPI gene have been observed in human red blood cells or mouse-brain capillary endothelial cells. Furthermore, the Alternative Splicing Database predicts three alternative splice variants for human TPI, but none of them has been identified experimentally to date. Along these lines, protein diversity is often generated by utilizing alternative translation Chlorhexidine hydrochloride initiation sites, mainly if a second start site is located within the first 20 codons. The usage of alternative translation initiation sites have been discussed for disease-causing proteins such as parkin, the prion protein and the breast cancer antigen BRCA1. Here, we provided evidence that the second in-frame ATG codon in the TPI gene encoding MET14 can be used as an alternative translation initiation site in yeast as well as in mammalian cells. However, the resulting protein did not suppress the growth defect of Dtpi1 yeast on glucose medium indicating that this variant lacks catalytic activity. This result is not unexpected, since this TPI variant lacks the catalytic lysine and at least three residues of the intersubunit interface. Interestingly, a TPI deficiency patient which has inherited a start codon mutation in combination with the mutation at position 104, has a stronger pathology in comparison to Glu104Asp homozygotes. Structural alterations of the pathogenic TPI variants have often been speculated to contribute to TPI deficiency as a number of mutations seem to affect the dimerization interface of TPI which forms a stable dimer in most investigated organisms. To address this issue, we have analyzed the dimerization properties between wild-type and the pathogenic TPI variants by a quantitative assay that is based on the lacZ gene as reporter. This allows determination of the relative strength of protein-protein interactions in yeast, however, these values can not be directly correlated with binding constants determined by in vitro measurements. We discovered that the dimerization behavior between wild-type TPI and the two pathogenic variants Cys41Tyr and Glu104Asp is strongly altered compared to the dimerization behavior between wild-type proteins. We further observed that the glycolytically inactive TPI2ndATG variant dimerizes with wild-type TPI, although at reduced levels in comparison to the dimerization of wild-type TPI proteins.
PCR was performed under conditions described earlier and all resultant products were verified by sequencing. As aforementioned, the frequency of TPI null alleles is much higher as the rare incidence of TPI deficiency, and in vitro measurements of a mutant TPI variant purified from E. coli demonstrated that the purified TPI protein carrying the Phe240Leu mutation exhibited a 6-fold higher activity than expected from the measurements performed with the patient erythrocyte extract. Moreover, theoretical calculations demonstrated that the extremely high DHAP level observed in patients’erythrocytes does not correlate with the measured residual TPI activity. Since patients’metabolites highly depend on the genetic background and environmental factors, we have generated an in vivo system Folinic acid calcium salt pentahydrate allowing analysis of the enzymatic activity of wild-type or pathogenic TPI variants inside a cell without additional side effects. At least three different isoelectric TPI variants encoded by the single TPI gene have been observed in human red blood cells or mouse-brain capillary endothelial cells. Furthermore, the Alternative Splicing Database predicts three alternative splice variants for human TPI, but none of them has been identified experimentally to date. Along these lines, protein diversity is often generated by utilizing alternative translation Chlorhexidine hydrochloride initiation sites, mainly if a second start site is located within the first 20 codons. The usage of alternative translation initiation sites have been discussed for disease-causing proteins such as parkin, the prion protein and the breast cancer antigen BRCA1. Here, we provided evidence that the second in-frame ATG codon in the TPI gene encoding MET14 can be used as an alternative translation initiation site in yeast as well as in mammalian cells. However, the resulting protein did not suppress the growth defect of Dtpi1 yeast on glucose medium indicating that this variant lacks catalytic activity. This result is not unexpected, since this TPI variant lacks the catalytic lysine and at least three residues of the intersubunit interface. Interestingly, a TPI deficiency patient which has inherited a start codon mutation in combination with the mutation at position 104, has a stronger pathology in comparison to Glu104Asp homozygotes. Structural alterations of the pathogenic TPI variants have often been speculated to contribute to TPI deficiency as a number of mutations seem to affect the dimerization interface of TPI which forms a stable dimer in most investigated organisms. To address this issue, we have analyzed the dimerization properties between wild-type and the pathogenic TPI variants by a quantitative assay that is based on the lacZ gene as reporter. This allows determination of the relative strength of protein-protein interactions in yeast, however, these values can not be directly correlated with binding constants determined by in vitro measurements. We discovered that the dimerization behavior between wild-type TPI and the two pathogenic variants Cys41Tyr and Glu104Asp is strongly altered compared to the dimerization behavior between wild-type proteins. We further observed that the glycolytically inactive TPI2ndATG variant dimerizes with wild-type TPI, although at reduced levels in comparison to the dimerization of wild-type TPI proteins.
In the present study we examined the effect of Vpu inactivation on Gag-Env interaction particle release
The clones were screened using Southern Blot analysis with a lentiviral specific probe and clones with a single integration were further investigated. Whereas almost no survival of undifferentiated cells was obtained for differentiated single integrant clones OT #4 and OT# 11, more than 1% of Oct-3/4 positive cells were still observed after GCV treatment of NT clones. This incomplete elimination might be due to an epigenetic silencing of the transgene. Another explanation for the presence of Oct-3/4 positive cells after GCV treatment and differentiation might be contaminations by a few untransduced ES cells. To avoid this, the single integrant clones can be further subcloned to achieve pure single integrant ES cell clones but this is more time consuming. Furthermore, in contrast to the generation of single clones, the usage of mixed ES cell populations is much simpler to handle since the additional process of clone selection and screening by Southern blot is not necessary. Irrespectively whether the TK vector is not functional or not present, a pre-selection strategy can be employed to obtain pure ES cell populations that express TK. Therefore, we incorporated hygromycin resistance gene as a pre-selection tool in the LVs to select for transduced ES cells. And indeed, treatment of STPH-transduced ES cells with hygromycin and GCV abolished undifferentiated cells in vitro. Importantly, no teratoma formation in the in vivo studies could be observed when mice were injected with these pre-selected transduced ES cells and treated with GCV. This could be a promising application of LVs and the TK/GCV system in future clinical approaches. The GCV treatment has to be applied directly after ES cell injection to prevent tumor formation caused by pluripotent cells as it is not able to eliminate or stop the growth of established tumors. This in accordance with previously published results. In addition, we tried to raise copy numbers of STPHtransduced ES cells to decline the untransduced cell population. Even with higher average copy numbers, several ES cells seemed to be not transduced because undifferentiated cells survived the GCV treatment. In conclusion, complete elimination was only achieved by hygromycin pre-selection. The functional interaction of Vpu with Gag through a tetratricopeptide repeat protein, UBP has been shown and its role as an intermediate between Vpu and Gag was proposed to play an important role in virus assembly. A recent study demonstrated that Vpu enhances virion release by preventing endosomal accumulation of Gag. In addition to its effect on Gag trafficking, Vpu slows down the progress of Env glycoproteins along biosynthetic pathways, from ER and Golgi to plasma membrane. On the other hand, Gag MA has been shown to have predominant effect on Envelope function. Previous studies demonstrated that specific amino acid substitutions in the Gag MA region abrogate incorporation of HIV-1 Env glycoproteins into virus particles and redirects Echinatin particle assembly to intracellular sites. Several studies suggest that the cytoplasmic domain of Env gp41 and the MA domain of Gag, together with various cellular cofactors, play a central role in HIV1 assembly and specific alterations in the MA region of Gag affects Env incorporation onto virion particles, association with lipid rafts 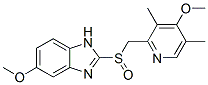 and thereby virion infectivity. While MA was shown to modulate Env assembly on mature virions, absence of Vpu has been shown to modulate Gag trafficking, assembly and infectivity besides modulation of particle release. Taken together, available information suggests interplay between Gag, Env and Vpu proteins and their Ginsenoside-Ro cross-talk seems to be important for assembly and production of infectious virus particles.
and thereby virion infectivity. While MA was shown to modulate Env assembly on mature virions, absence of Vpu has been shown to modulate Gag trafficking, assembly and infectivity besides modulation of particle release. Taken together, available information suggests interplay between Gag, Env and Vpu proteins and their Ginsenoside-Ro cross-talk seems to be important for assembly and production of infectious virus particles.
It is assumed that one of the cuticle’s functions is to protect the plant surface from possible
In their study, the authors also detected the increased expression of two ER stress related transcription factors, namely CAAT enhancer binding protein b and growth arrest and DNA-damage inducible protein 153 also known as C/EBPhomologous protein. Interestingly, a recent study showed that stress-induced expression of CHOP was associated with repression MyoD, a gene involved in muscle differentiation. The murine dnajb3 promoter has two C/EBP binding sites and their role in the regulation of DNAJB3 following activation of the ER stress, if any, remains to be demonstrated. Further clarifications are also needed to confirm whether or not the observed inhibition of DNAJB3 in vitro following activation of the ER stress with palmitate occurs also in vivo in obese subjects. In our study population, we detected high levels of CHOP as well as the spliced form of the ER stress response protein, X-box-binding protein 1 in obese subjects which indicates that the ER stress is induced in obese subjects. On the other 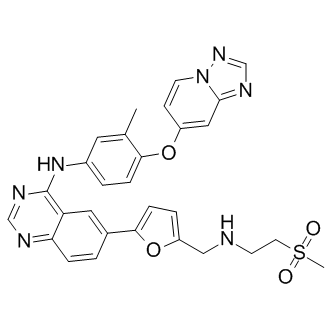 hand, our data did not rule out the effect of other inflammatory mediators since we only selected in our study a few representatives of inflammatory cytokines. Another limitation of the study is the fact that obese subjects that participated in this study are relatively older than lean subjects. In this regard, aging is well known to decrease the expression of HSPs such as HSP-72. However, the fact that, in one hand, the expression of HSC-70 and HSP-90 were increased in obese subjects and on the other hand, the positive effect of exercise in restoring the normal expression of DNAJB3 suggest that aging is unlikely to affect the expression of DNAJB3. In conclusion, we provided compelling evidence that the expression of the co-chaperone DNAJB3 is markedly reduced at the mRNA and protein levels in both PBMC and adipose tissue of obese subjects. We further demonstrated that physical exercise restores the normal expression of DNAJ3 to the levels comparable to lean subjects. Although we did not demonstrate the causal relationship between reduced expression of DNAJB3 and obesity, we demonstrated that DNAJB3 is part of a complex that contains key proteins involved in obesity, Atropine sulfate insulin resistance and T2D such as HSP-72, JNK and IKKb. All together, our data support the suggestion that DNAJB3 can potentially play a protective role against obesity, and thus targeting DNAJB3 may have a potential therapeutic benefit for the control and management of obesity and insulin resistance. Plants encounter a wide range of pathogens and insects in their natural environment. Some of these are responsible for annual worldwide economic damage due to losses in important agricultural crops. Throughout their coevolution with pathogens, plants have developed both physical barriers and physiological responses, which aid them in Ginsenoside-Ro coping with pathogen attacks. The first barrier protecting plants from pathogens is a physical one�Cthe cuticle, which is defined as a noncellular waxy structure that covers the epidermal cells. The cuticular layer covers all of the aerial organs and plays multiple roles in plants, including regulation of epidermal permeability and nonstomatal water loss, and protection against insects, pathogens, UV light and frost. The second barrier protecting plants from pathogenic attack is a set of biochemical reactions, which lead to hypersensitive and acquired immune responses. These constitutive and inducible defense events depend largely on the perception of signaling molecules, some of which can be activated by cuticle-degradation products. It is generally accepted that the cuticle’s mechanical strength is provided by the cutin matrix, a polymer formed by threedimensional crosslinking of covalent bonds.
hand, our data did not rule out the effect of other inflammatory mediators since we only selected in our study a few representatives of inflammatory cytokines. Another limitation of the study is the fact that obese subjects that participated in this study are relatively older than lean subjects. In this regard, aging is well known to decrease the expression of HSPs such as HSP-72. However, the fact that, in one hand, the expression of HSC-70 and HSP-90 were increased in obese subjects and on the other hand, the positive effect of exercise in restoring the normal expression of DNAJB3 suggest that aging is unlikely to affect the expression of DNAJB3. In conclusion, we provided compelling evidence that the expression of the co-chaperone DNAJB3 is markedly reduced at the mRNA and protein levels in both PBMC and adipose tissue of obese subjects. We further demonstrated that physical exercise restores the normal expression of DNAJ3 to the levels comparable to lean subjects. Although we did not demonstrate the causal relationship between reduced expression of DNAJB3 and obesity, we demonstrated that DNAJB3 is part of a complex that contains key proteins involved in obesity, Atropine sulfate insulin resistance and T2D such as HSP-72, JNK and IKKb. All together, our data support the suggestion that DNAJB3 can potentially play a protective role against obesity, and thus targeting DNAJB3 may have a potential therapeutic benefit for the control and management of obesity and insulin resistance. Plants encounter a wide range of pathogens and insects in their natural environment. Some of these are responsible for annual worldwide economic damage due to losses in important agricultural crops. Throughout their coevolution with pathogens, plants have developed both physical barriers and physiological responses, which aid them in Ginsenoside-Ro coping with pathogen attacks. The first barrier protecting plants from pathogens is a physical one�Cthe cuticle, which is defined as a noncellular waxy structure that covers the epidermal cells. The cuticular layer covers all of the aerial organs and plays multiple roles in plants, including regulation of epidermal permeability and nonstomatal water loss, and protection against insects, pathogens, UV light and frost. The second barrier protecting plants from pathogenic attack is a set of biochemical reactions, which lead to hypersensitive and acquired immune responses. These constitutive and inducible defense events depend largely on the perception of signaling molecules, some of which can be activated by cuticle-degradation products. It is generally accepted that the cuticle’s mechanical strength is provided by the cutin matrix, a polymer formed by threedimensional crosslinking of covalent bonds.
These six protein clusters contained 149 candidate Parkin-binding proteins in total
Whereas most of the candidates are functionally similar amongst themselves, in cluster 6 the candidates are all functionally related to the PD-related proteins, with the only exception of YWHAE. A sub-network containing only the candidate proteins that are functionally similar to the PD-related proteins includes 37 proteins in total and 29 candidate Parkin-binding proteins. Twenty-eight candidates fall into four clusters of enriched GO processes that cover the functions of the known PD proteins, including the various known functions of Parkin. Among these processes we have identified several proteins involved in cell death: HSPD1, CASP14, H1F0, DAP3, AIFM1, GSDMA, SET, OPA1, and BAG2. In addition, TOMM70A, DAP3 and OPA1 are involved in mitochondrion organization, and CCT3, CLPX1, HSPD1, PRKCSH and BAG2 are related to protein folding. Of the 29 candidates that are functionally similar to the PD-related proteins, 13 were also identified in a genetic screen as modifiers of Parkin and PINK1 mutant functions in Drosophila, providing further evidence for the biological relevance of the interactions. The diverse functions of the Parkin protein partners reported here are consistent with the functional diversity of the pathogenic processes associated with Parkin-linked parkinsonism. Parkin is localized in the endoplasmic reticulum, the Golgi apparatus, the outer nuclear membrane, synaptic vesicles, and the outer LOUREIRIN-B mitochondrial membrane, and there is a large body of evidence showing that Parkin can interfere with a diverse range of cellular processes and pathways. Like other E3 ubiquitin ligases, it is a component of the ubiquitin proteasome system, a main cellular 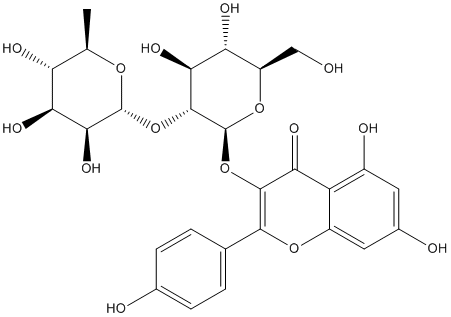 pathway that promotes removal of damaged or misfolded proteins, it is involved in signal transduction, protein and membrane trafficking and transcriptional regulation, replication and transcription of mitochondrial DNA, mitophagy, neuroprotection, and apoptosis. Parkin expression has been reported to protect cells against multiple forms of stress, but although the exact mechanism of this prosurvival function remains elusive, accumulating evidence exists that it involves inhibition of programmed cell death. Two recent studies identified Bax and the mitochondrial pro-apoptotic protein ARTS as Parkin substrates that both might contribute to the anti-apoptotic effect of Parkin. In our study, we identified novel associations between Parkin and several proteins involved in cell death processes. An interaction of Parkin with one of them, OPA1, is supported by the observation that inactivation of OPA1, which promotes mitochondrial fusion, rescues the phenotypes of cell death, muscle degeneration, and mitochondrial abnormalities in Parkin and PINK1 mutants in Drosophila. DAP3, another candidate protein involved in cell death, mediates mitochondrial fragmentation, probably reflecting its role in mitochondrial fission. Both proteins might have a role in cell death-associated changes in mitochondrial morphology mediated by Parkin. TOMM70A, which encodes a component of a translocase complex of the outer mitochondrial membrane involved in the import of mitochondrial precursor proteins, has been associated recently to Parkin as it is degraded by the UPS after translocation of Parkin to mitochondria. LRPPRC, which was identified already in a proteomic Diperodon analysis of Parkin interactors, might be involved in mitophagic initiation, maturation, trafficking, and lysosomal clearance through its interaction with the MAP1S protein. Other Parkin-binding proteins, like HSPD1 and CLPX, are involved in protein folding and response to unfolded proteins.
pathway that promotes removal of damaged or misfolded proteins, it is involved in signal transduction, protein and membrane trafficking and transcriptional regulation, replication and transcription of mitochondrial DNA, mitophagy, neuroprotection, and apoptosis. Parkin expression has been reported to protect cells against multiple forms of stress, but although the exact mechanism of this prosurvival function remains elusive, accumulating evidence exists that it involves inhibition of programmed cell death. Two recent studies identified Bax and the mitochondrial pro-apoptotic protein ARTS as Parkin substrates that both might contribute to the anti-apoptotic effect of Parkin. In our study, we identified novel associations between Parkin and several proteins involved in cell death processes. An interaction of Parkin with one of them, OPA1, is supported by the observation that inactivation of OPA1, which promotes mitochondrial fusion, rescues the phenotypes of cell death, muscle degeneration, and mitochondrial abnormalities in Parkin and PINK1 mutants in Drosophila. DAP3, another candidate protein involved in cell death, mediates mitochondrial fragmentation, probably reflecting its role in mitochondrial fission. Both proteins might have a role in cell death-associated changes in mitochondrial morphology mediated by Parkin. TOMM70A, which encodes a component of a translocase complex of the outer mitochondrial membrane involved in the import of mitochondrial precursor proteins, has been associated recently to Parkin as it is degraded by the UPS after translocation of Parkin to mitochondria. LRPPRC, which was identified already in a proteomic Diperodon analysis of Parkin interactors, might be involved in mitophagic initiation, maturation, trafficking, and lysosomal clearance through its interaction with the MAP1S protein. Other Parkin-binding proteins, like HSPD1 and CLPX, are involved in protein folding and response to unfolded proteins.