Taken together, this study demonstrates that miR-34a may be required for the anti-cancer effects of DADS, both in human breast cancer cell lines and in xenograft models. Furthermore, SRC may be a critical target of miR-34a. DADS may therefore suppress proliferation and invasion in human breast cancer cells by up-regulating miR-34a and consequently reducing SRC/Ras/ ERK signaling, indicating that miR-34a may serve as a potential gene therapy target for enhancing the antitumor effects of DADS in breast cancer therapy. MicroRNAs, a class of small non-coding RNA, are widely studied as crucial regulatory molecules able to modulate broad regulatory networks at the post-transcriptional levels. miRNA is generated from primary miRNA and precursor miRNA in animals, with the premiRNA presenting a stable stem-loop structure. Both of the two arm products, miR5p and miR3p, have been reported to form mature and functional miRNAs, with the loop sequences connecting miR5p and miR3p in the stem-loop structure typically degraded during miRNA biogenesis. These transitory intermediates are rarely examined due to an unseen direct role in the miRNA regulation process. Instead, most studies have focused on the functional and regulatory roles of miRNAs during miRNA-mRNA recognition as it relates to expression or translational repression. However, indeed, both pri-miRNA and pre-miRNA may contribute to the regulatory process. Specifically, the loop nucleotides may tune and alter miRNA activity, controlling the processing precision during the miRNA maturation process. Recently, the potential impact of hairpin loops has been examined, with short hairpin RNA loops possibly influencing effectivity and alternative loop conformations in miRNAs potentially effecting expression. The loop sequence may also affect Dicer recognition and possibly specificity, thus affect miRNA cleavage during the maturation process. As mentioned above, the loop sequence in premiRNAs has an important role in regulating the activities and specificities of related molecules that can facilitate Clofentezine Drosha and Dicer, with a mutation in the loop possibly affecting miRNA processing. Furthermore, miRNAs have been a key focal point because a miRNA locus can yield multiple sequences have diverse 59 and/or 39 ends and expression levels. These miRNA variants, or physiological miRNA isoforms, may be mainly generated by imprecise Drosha and Dicer cleavage during pre-miRNA processing. Additionally, the loop sequences may also impact pre-miRNA processing by affecting Drosha and/or Dicer directly or indirectly. Even though premiRNA loop sequences have been shown to directly or indirectly impact Drosha and Dicer during miRNA processing, few studies have performed an evolutionary analysis of the loop sequences, especially one focused on homology and clustered 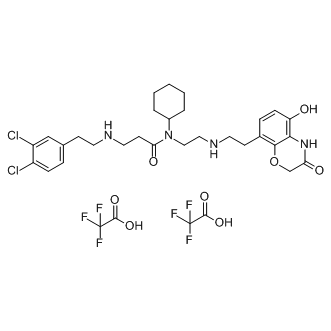 miRNAs across Atractylenolide-III different animal species. Herein, we attempt to perform an evolutionary analysis of the loop sequences in let-7 and locate related miRNAs across different animal species. The let-7 gene family has been widely detected in metazoans, with its associated miRNAs thought to have coexpanded with the HOX gene clusters. A series of homologous miRNAs can be found in a miRNA gene family, including multicopy pre-miRNAs from specific animal species. Simultaneously, some members can be located in a cluster with other related miRNAs, with miRNAs within a close proximity on a chromosome being co-transcribed as a polycistronic transcript. Therefore, a classical miRNA gene family and related miRNAs are typically selected to track and reveal the evolutionary patterns of the ever present yet ignored loop sequences. Thus, this study will examine the roles of loops during miRNA maturation and processing.
miRNAs across Atractylenolide-III different animal species. Herein, we attempt to perform an evolutionary analysis of the loop sequences in let-7 and locate related miRNAs across different animal species. The let-7 gene family has been widely detected in metazoans, with its associated miRNAs thought to have coexpanded with the HOX gene clusters. A series of homologous miRNAs can be found in a miRNA gene family, including multicopy pre-miRNAs from specific animal species. Simultaneously, some members can be located in a cluster with other related miRNAs, with miRNAs within a close proximity on a chromosome being co-transcribed as a polycistronic transcript. Therefore, a classical miRNA gene family and related miRNAs are typically selected to track and reveal the evolutionary patterns of the ever present yet ignored loop sequences. Thus, this study will examine the roles of loops during miRNA maturation and processing.
Monthly Archives: May 2019
The rapid clearance upon reinfection and involvement of Th17 and Th1 responses
Translation of biomarkers from mouse to man however should be performed with care. Overall, our study describes the molecular and cellular sequence of events that eventually lead to the induction of protective immunity to pertussis, including local immunity in the lungs, which is not induced by current vaccines. Especially the inductions of Th1 and Th17 cells and IgA in the lungs are regarded key elements in superior immunity. Our data can be used to select vaccine concepts that resemble infection, with respect to e.g. administration route and prolonged antigen exposure, and immune signatures elicited. Local delivery of vaccines may be an important advancement for improved protection as was previously demonstrated with a live attenuated pertussis vaccine. Also adjuvants could be selected and optimized that can steer host immunity of inactivated vaccines towards an infection-like immune response with minimal adverse effects. The molecular and cellular fingerprint of the local immune response discovered in B. pertussis Ginsenoside-F4 infected mice thus can provide a strong guidance for developing improved pertussis vaccines. KLF4 has been 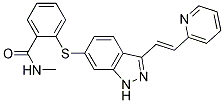 shown to regulate cell proliferation and differentiation, its role has been extensively investigated in several human cancers by using gain- and loss-of-function approaches. In colon and prostate cancer, KLF4 acts as an oncogene. In contrast, it plays a tumor suppressor role in neuroblastoma, lung cancer, gastric cancer, lymphoma, cervical cancer, pancreatic ductal cancer, and Dimesna hepatocellular carcinoma. In breast cancer, KLF4 can function both as an oncogene and a tumor suppressor. The role of KLF4 in ovarian cancer has not been adequately and mechanistically addressed. A previous study indicates that the expression level of KLF4 was significantly reduced in ovarian cancer compared to normal ovarian epithelium, suggesting that KLF4 might potentially act as a tumor suppressor in ovarian cancer. KLF4 plays a unique role in stem cell reprogramming by facilitating the mesenchymal to epithelial transition. The cellular phenotypic switch from epithelial to mesenchymal cell transition is a fundamental process in tumor metastasis that is a prominent feature of ovarian carcinomas. The MET or EMT leads to the alterations of epithelial and mesenchymal marker gene expression that include snail1 & 2, Zeb 1 &2, Twist, vimentin, Ecadherin. EMT is regulated by multiple signaling pathways, which include WNT, TGFb, and Notch.. Recent studies indicate that miRNAs regulate EMT or MET pathways by targeting epithelial or mesenchymal cell marker genes that include miR-194, miR-203, and miR-200c. KLF4 has been shown to regulate EMT in several different cancer cells. In hepatocellular carcinoma, breast, and prostate cancer cells, KLF4 activates the transcription of the epithelial cell marker gene Ecadherin and represses the mesenchymal cell marker gene snail 2 by binding to their respective promoters. KLF4 in these cancers promotes MET and inhibits tumor cell growth. In the present study, we investigated the role of KLF4 in ovarian cancer cells using a doxycycline -dependent KLF4-inducible lentiviral vector and found that inducible overexpression of KLF4 reduced cell proliferation, migration, and invasion through promoting MET in ovarian cancer cells. In this study, we investigated the role of KLF4 in ovarian cancer cells using lentiviral vector mediated inducible expression.
shown to regulate cell proliferation and differentiation, its role has been extensively investigated in several human cancers by using gain- and loss-of-function approaches. In colon and prostate cancer, KLF4 acts as an oncogene. In contrast, it plays a tumor suppressor role in neuroblastoma, lung cancer, gastric cancer, lymphoma, cervical cancer, pancreatic ductal cancer, and Dimesna hepatocellular carcinoma. In breast cancer, KLF4 can function both as an oncogene and a tumor suppressor. The role of KLF4 in ovarian cancer has not been adequately and mechanistically addressed. A previous study indicates that the expression level of KLF4 was significantly reduced in ovarian cancer compared to normal ovarian epithelium, suggesting that KLF4 might potentially act as a tumor suppressor in ovarian cancer. KLF4 plays a unique role in stem cell reprogramming by facilitating the mesenchymal to epithelial transition. The cellular phenotypic switch from epithelial to mesenchymal cell transition is a fundamental process in tumor metastasis that is a prominent feature of ovarian carcinomas. The MET or EMT leads to the alterations of epithelial and mesenchymal marker gene expression that include snail1 & 2, Zeb 1 &2, Twist, vimentin, Ecadherin. EMT is regulated by multiple signaling pathways, which include WNT, TGFb, and Notch.. Recent studies indicate that miRNAs regulate EMT or MET pathways by targeting epithelial or mesenchymal cell marker genes that include miR-194, miR-203, and miR-200c. KLF4 has been shown to regulate EMT in several different cancer cells. In hepatocellular carcinoma, breast, and prostate cancer cells, KLF4 activates the transcription of the epithelial cell marker gene Ecadherin and represses the mesenchymal cell marker gene snail 2 by binding to their respective promoters. KLF4 in these cancers promotes MET and inhibits tumor cell growth. In the present study, we investigated the role of KLF4 in ovarian cancer cells using a doxycycline -dependent KLF4-inducible lentiviral vector and found that inducible overexpression of KLF4 reduced cell proliferation, migration, and invasion through promoting MET in ovarian cancer cells. In this study, we investigated the role of KLF4 in ovarian cancer cells using lentiviral vector mediated inducible expression.
Whether RGZ alters hepatic Sirt1 and to investigate the potential synergistic effects
Despite some interesting findings in this study, our analyses had a limitation that part of the data regarding the involvement of Sirt1/6 in the RGZ’s effects was based on the alteration of mRNA expression. In general, the mRNA expression of specific target gene corresponds to its protein levels. However, it is possible to have a discrepancy between mRNA expression and protein levels due to post-transcriptional, translational or post-translational regulations. The present study was conducted to elucidate of Sirt1 and Sirt6 on regulation of hepatic fat accumulation by RGZ. Using a rat model of moderate obesity and insulin resistance and a cell model of hepatocyte steatosis, we report upregulation of adiponectin, Sirt1/6, and downstream D-Pantothenic acid sodium targets of Sirt1/6, and also, increases in both LKB1 and AMPK activities following RGZ treatment. Additionally, we demonstrated that Sirt1/6 knockdown abolished the effects of RGZ with regard to hepatocyte fat accumulation and the Sirt1/6-AMPK pathway, suggesting that Sirt1/6 regulates RGZ-mediated metabolic processes and that the relationship between Sirt 1 and Sirt6 may be compensatory without synergistic effects. These findings indicate that Sirt1 and Sirt6 are involved in the RGZ-mediated improvement of hepatocyte steatosis, and warrant future study to identify the interaction of different sirtuins on metabolic regulation. Glaucoma is a significant cause of irreversible blindness worldwide. Primary open angle glaucoma is the most common type. Older age, a family history of POAG, myopia, central corneal thickness, and ocular hypertension are relatively consistent risk factors for POAG. Diabetes mellitus is a serious and increasingly prevalent health problem worldwide due to lifestyle changes and an aging population. DM is associated with severe acute and chronic complications, which negatively influence both the quality of life and the survival of affected individuals. Some Butenafine hydrochloride studies also found that DM is another possible risk factor for POAG. However, the relationship between DM and POAG is controversial. An earlier meta-analysis in 2004 indicated that DM is associated with an increased risk of developing POAG. However, that meta-analysis of seven cross-sectional and five case-control studies of DM and the risk of POAG did not include any cohort studies. The major drawback of cross-sectional studies is that they cannot establish clear temporal relationship between exposure and outcome. Prospective cohort studies would be a good way to assess the relationship between exposure to diabetes and development of POAG. Since then, much larger-scale epidemiological evidence, especially cohort studies, of an association between DM and POAG has been reported. However, these epidemiological studies of the relationship between DM and POAG were somewhat contradictory and inconclusive, and two studies reported discordant results. To provide a more accurate estimate of the association between DM and POAG, we conducted a meta-analysis of all case-control and cohort studies involving DM and POAG. Although several risk factors for the development of POAG have been evaluated, this is a field of ongoing investigation. DM is a possible 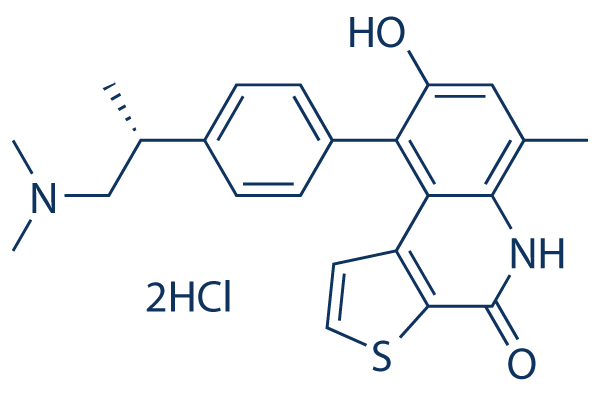 risk factor for POAG. The current literature does not provide a definitive link between DM and POAG. Hence, the purpose of this systematic review and meta-analysis was to summarize all the available relevant evidence with reference to the relationship between DM and POAG.
risk factor for POAG. The current literature does not provide a definitive link between DM and POAG. Hence, the purpose of this systematic review and meta-analysis was to summarize all the available relevant evidence with reference to the relationship between DM and POAG.
This is the first evidence showing the RGZ-mediated alteration of hepatic Sirt1 in a rodent model of NAFLD
In previous reports from our group, the protective action of RGZ against hepatic steatosis is by coordinated regulation of adiponectin, Sirt6, and AMPK. Up-regulation of Sirt1/6 and the downstream target genes of Sirt1/6 as well as activation of LKB1/AMPK by RGZ led us to hypothesize that RGZ may exert its positive Diacerein effects by acting on both Sirt1 and Sirt6. From these observations and accumulating evidence of sirtuins and their metabolic regulation, we speculate that Sirt1 and Sirt6 possess similarities in cellular localization and metabolic functions and question whether both sirtuins are synergistic or compensatory. To determine whether Sirt1 and Sirt6 are causally related to the development of hepatic steatosis and its amelioration by RGZ, and whether their actions are synergistic, we directly inhibited the Sirt1/6 pathway using RNAi-mediated gene silencing targeting Sirt1 and/or Sirt6 in AML12 mouse hepatocytes. Sirt1/6 knockdown aggravated hepatocyte fat accumulation as shown by increased TG content and suppressed the favorable effects of RGZ on hepatocyte steatosis. In addition, Sirt1/6 knockdown suppressed gene expression of Ppargc1a and Foxo1 as expected, and abolished the RGZ-mediated activation of LBK1 and AMPK. However, double knockdown of Sirt1 and Sirt6 did not result in a synergistic effect on the RGZ-mediated alterations. Conventionally, it has been considered that Sirt1 and Sirt6 exert their functions as NAD -dependent deacetylase and ADP-ribosyltransferase. However, provocative reports have suggested that Sirt1 may be responsible for transcriptional regulation of downstream target genes. Sirt1 upregulated adiponectin gene expression in fully differentiated 3T3-L1 adipocytes by enhancing Foxo1 and C/enhancer-binding protein alpha interaction with the adiponectin promoter. A positive correlation between Sirt1 and Sirt6 has been reported in mice, and, additionally, the increase in Sirt1 occurred earlier than that of Sirt6 in the mouse liver during fasting. Additionally, whether Sirt1 is responsible for transcriptional regulation of Sirt6 was ![]() investigated, and the results showed that Sirt1 induced Sirt6 gene expression by forming a complex with Foxo3a and nuclear respiratory factor 1-binding sites on the Sirt6 promoter. Based on these observations, we speculated that Sirt1 and Sirt6 may have synergistic effects on metabolic regulation. However, no synergistic effect between Sirt1 and Sirt6 was observed in this study, suggesting that the relationship between Sirt1 and Sirt6 may be compensatory as a back-up system. Although we could not demonstrate the synergistic effects of two isoforms of sirtuins, our results suggest the possible interaction between Sirt1 and Sirt6. In normal AMP12 hepatocytes without PA 3,4,5-Trimethoxyphenylacetic acid treatment, Sirt1 mRNA expression was not altered by Sirt6 knockdown, but, Sirt6 mRNA expression tends to be lowered by Sirt1 knockdown. These results support the previous findings showing that Sirt1 is responsible for transcriptional regulation of Sirt6. In addition, regarding the effects on Ppargc1a and Foxo1, Sirt1 knockdown resulted in more profound suppression than Sirt6 knockdown. In a hepatocyte steatosis model with PA treatment, the regulatory effects of RGZ on Sirt1 were significantly blunted by Sirt6 knockdown, which suggests that upregulation of Sirt1 by RGZ was partially dependent on Sirt6. On the other hand, RGZ’s effects on Sirt6 were not altered by Sirt1 knockdown in a hepatocyte steatosis model.
investigated, and the results showed that Sirt1 induced Sirt6 gene expression by forming a complex with Foxo3a and nuclear respiratory factor 1-binding sites on the Sirt6 promoter. Based on these observations, we speculated that Sirt1 and Sirt6 may have synergistic effects on metabolic regulation. However, no synergistic effect between Sirt1 and Sirt6 was observed in this study, suggesting that the relationship between Sirt1 and Sirt6 may be compensatory as a back-up system. Although we could not demonstrate the synergistic effects of two isoforms of sirtuins, our results suggest the possible interaction between Sirt1 and Sirt6. In normal AMP12 hepatocytes without PA 3,4,5-Trimethoxyphenylacetic acid treatment, Sirt1 mRNA expression was not altered by Sirt6 knockdown, but, Sirt6 mRNA expression tends to be lowered by Sirt1 knockdown. These results support the previous findings showing that Sirt1 is responsible for transcriptional regulation of Sirt6. In addition, regarding the effects on Ppargc1a and Foxo1, Sirt1 knockdown resulted in more profound suppression than Sirt6 knockdown. In a hepatocyte steatosis model with PA treatment, the regulatory effects of RGZ on Sirt1 were significantly blunted by Sirt6 knockdown, which suggests that upregulation of Sirt1 by RGZ was partially dependent on Sirt6. On the other hand, RGZ’s effects on Sirt6 were not altered by Sirt1 knockdown in a hepatocyte steatosis model.
Significant increases of both circulating granulocytes and monocytes could be demonstrated in recognize several endogenous proteins
When released upon cellular stress or injury such as Sipeimine during MI. Toll-like receptors constitute one of the largest subfamilies of PRRs. Of these, TLR9 has been demonstrated to specifically detect unmethylated DNA, rich in cytosine-phosphate-guanine motifs. Recent work has identified mitochondrial DNA to function as a DAMP, causing activation of the innate immune system through TLR9. On this notion, our group have previously reported increased circulating levels of mtDNA upon PCI of human MI. The consequence of TLR signaling in I/R remains unclear. Activation of TLR9 prior to I/R lead to reduced infarct extension and improved cardiac function. From a clinical point of view, an important and hereto unaddressed question is whether activation of said receptors during onset of I/R also impacts subsequent myocardial damage. Thus, in the current study we investigated the pathophysiological consequence of intervening with the TLR9-agonist CpG B during onset of ischemia. Evaluation of infarct size was performed as previously described. Briefly, after 24 hours of reperfusion, mice were anesthetized, the thoracic cavity opened and the slipknot re-ligated. The aorta ascendens was subsequently clamped and approximately 0.2 ml 2% Evans blue dye was injected into the proximal portion of the aorta, this allowing coronary perfusion and visualization of the area at risk. Hearts were subsequently excised, let to freeze on dry-ice before cross-sectioned into standardized 1 mm slices by the use of a mouse heart slicer matrix. The main objective of this study was to investigate the effect of TLR9-activation during onset of myocardial I/R with respect to infarct size. Although TLR9-activation elicited robust increases in systemic inflammation as well as a decrease in some inflammatory markers within the myocardium, these systemic and cardiac inflammatory alterations did not translate into changes in infarct size. Several previous reports have convincingly demonstrated beneficial effects of TLR-stimulation, including TLR9, initiated before ischemia. These studies do advocate for a putative role of TLR-activation in clinical situations of expected cardiac ischemia. 4-(Benzyloxy)phenol However, no studies have investigated the effects of TLR-stimulation restricted to the more common clinical scenario; i.e. intervention upon ischemia/reperfusion. Even though still clinically relevant, our experimental setup does not allow discriminating the effects of our intervention on ischemia, reperfusion or a combination of both. In order to specifically address the reperfusion phase of I/R, drugs should preferably be injected directly into the occluded artery upon reperfusion. As to our knowledge, this is not feasible in a murine I/ R-model. However, a requirement for substantial cardiac influences during ischemia necessitates drugs to enter deep into the myocardium at risk as well as temporal accessibility within the time-range of ischemia. As to the latter, we were not able to detect increased TLR9-signaling at earlier time-points than 30 minutes post-injection. Thus, even though we cannot exclude cardiac influences during ischemia, our experimental set-up suggests a predominant effect of TLR9-stimulation during reperfusion. Our main parameter, i.e. infarct extension at 24 hours, was similar between TLR9 stimulated and vehicle treated I/R mice However, we do provide evidence of substantial systemic inflammatory responses upon i.p. injection of CpG B. After CpG B 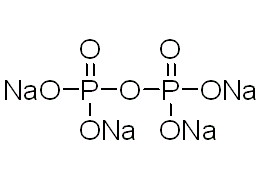 injection.
injection.