QSAR and molecular docking are integrated for the first time with experimental bioassay to guide rational design of mosquito repellents in a way similar to that widely used in drug discovery. The potential of the proposed integrated approach first tested on Aedes aegypti repellents will be applied in the future to the development of a broader class of vector control systems. Normalized and weighted values of the molecular fields are mapped on the molecular supergraph, with different shades of red and blue representing varying contributions, or Z-VAD-FMK impacts. These colorcoded schemes are given in Figure 5 for the atomic charge, van der Waals radii, hydrogen bond acceptor, hydrogen bond donor, and lipophilicity descriptors. Positions color-marked in more than one molecular field are the most informative ones. For example, a decrease in atomic charge and hydrophobicity coupled with an increase in van der Waals radius of the atom in position 2 leads to an increase in the antilog MED value. As it follows 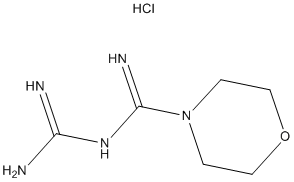 from the descriptor values given in Table 3, for the atoms occupying position 2, the terminal CH3 group is preferable both over the -CH2- group and a vacancy. This can be found useful for inferring an optimal length of alkyl chains. Similar analysis and conclusions can be made for position 86. The mosquito olfactory mechanism is complex, and OBP1 is most likely just the first step in developing a response. OBP1 either brings the odorant molecule into a direct contact with the olfactory receptors or exerts an allosteric action upon the OR. In both cases, the initial binding state of the AaegOBP1-odorant complex can change dramatically as the recognition event proceeds. It is quite reasonable to assume that the compounds sharing the 2-phenylcyclohexanol scaffold are bound in such a favorable mode that they are able to activate the OR machinery. FGF21 is expressed in multiple metabolic tissues. Moreover, FGF21 increases glucose uptake and insulin sensitivity while decreasing gluconeogenesis, lipogenesis and circulating cholesterol levels in multiple pre-clinical animal models when administered pharmacologically. Fasting and ketogenic conditions induce hepatic expression of FGF21 through PPAR��, while its expression is induced in adipose tissues by PPAR�� activation. FGF21 binds to fibroblast growth factor receptors complexed with the co-receptor ��Klotho and signals through phosphorylation cascades involving FRS2a and Erk/MAPK leading to transcriptional changes of several metabolic genes. Phosphorylation of Gsk-3, Shp2, Mek1/2, and Stat3 following FGF21 treatment of 3T3L1 adipocytes has been reported. There is growing evidence suggesting that FGF21 signaling specifically in adipose tissue is crucial for its beneficial metabolic effects. For example, FGFR1 and ��Klotho expression in adipose tissues, and downstream FGF21 signaling, are required for at least part of FGF21��s acute insulin-sensitizing and glucose uptake effects in mice. In support of this observation, FGF21 displays impaired glucose- and triglyceride-lowering efficacy in lipodystrophic mice. Recent reports have identified adiponectin as a key mediator of FGF21’s effects on glucose homeostasis and insulin sensitivity in mice. In addition, administration of FGF21 has been shown to induce weight loss and white adipose browning in several pre-clinical animal models. In order to further our VE-822 understanding of FGF21 signaling downstream of receptor activation in mouse adipose tissues, we have used unbiased transcriptomic and phosphoproteomic profiling to quantify transcripts and phosphoproteins that are modulated after acute FGF21 treatment. For our studies, we used both native FGF21 protein and a pegylated form that increases its half-life while displaying acute efficacy in terms of normalizing insulin-stimulated glucose uptake in insulin-resistant mice.
from the descriptor values given in Table 3, for the atoms occupying position 2, the terminal CH3 group is preferable both over the -CH2- group and a vacancy. This can be found useful for inferring an optimal length of alkyl chains. Similar analysis and conclusions can be made for position 86. The mosquito olfactory mechanism is complex, and OBP1 is most likely just the first step in developing a response. OBP1 either brings the odorant molecule into a direct contact with the olfactory receptors or exerts an allosteric action upon the OR. In both cases, the initial binding state of the AaegOBP1-odorant complex can change dramatically as the recognition event proceeds. It is quite reasonable to assume that the compounds sharing the 2-phenylcyclohexanol scaffold are bound in such a favorable mode that they are able to activate the OR machinery. FGF21 is expressed in multiple metabolic tissues. Moreover, FGF21 increases glucose uptake and insulin sensitivity while decreasing gluconeogenesis, lipogenesis and circulating cholesterol levels in multiple pre-clinical animal models when administered pharmacologically. Fasting and ketogenic conditions induce hepatic expression of FGF21 through PPAR��, while its expression is induced in adipose tissues by PPAR�� activation. FGF21 binds to fibroblast growth factor receptors complexed with the co-receptor ��Klotho and signals through phosphorylation cascades involving FRS2a and Erk/MAPK leading to transcriptional changes of several metabolic genes. Phosphorylation of Gsk-3, Shp2, Mek1/2, and Stat3 following FGF21 treatment of 3T3L1 adipocytes has been reported. There is growing evidence suggesting that FGF21 signaling specifically in adipose tissue is crucial for its beneficial metabolic effects. For example, FGFR1 and ��Klotho expression in adipose tissues, and downstream FGF21 signaling, are required for at least part of FGF21��s acute insulin-sensitizing and glucose uptake effects in mice. In support of this observation, FGF21 displays impaired glucose- and triglyceride-lowering efficacy in lipodystrophic mice. Recent reports have identified adiponectin as a key mediator of FGF21’s effects on glucose homeostasis and insulin sensitivity in mice. In addition, administration of FGF21 has been shown to induce weight loss and white adipose browning in several pre-clinical animal models. In order to further our VE-822 understanding of FGF21 signaling downstream of receptor activation in mouse adipose tissues, we have used unbiased transcriptomic and phosphoproteomic profiling to quantify transcripts and phosphoproteins that are modulated after acute FGF21 treatment. For our studies, we used both native FGF21 protein and a pegylated form that increases its half-life while displaying acute efficacy in terms of normalizing insulin-stimulated glucose uptake in insulin-resistant mice.
Monthly Archives: July 2019
This may be caused by failure in endodermal cell migration to the EB in the middle region of the embryos
Expression of Bmp4 in the lateral plate mesoderm, the extraembryonic mesoderm and the extraembryonic ectoderm was found in mutant embryos as in the wt. Tbx20 was expressed in the allantois, a derivative of the extraembryonic mesoderm, and in the linear heart tube, which derives from the cardiac subregion of the anterior lateral plate mesoderm in the wt. Again, both expression domains were established at opposite poles of the mutant embryos but neither domain was appropriately developed into an allantois. Finally, Gja, that is expressed in the cardiac endoderm and dorsal aorta in wt embryos was restricted to cell clusters at the anterior and posterior pole of the mutant embryos but clearly lacked an organization into vessel like structures. Together, this analysis suggests that PKCi-deficient embryos exhibit a normal anterior-posterior polarization of their main body axis. Mesoderm formation is initiated but comes to a premature halt, mesoderm differentiation into axial, paraxial, lateral and extraembryonic subtypes occurs 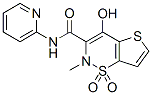 but subsequent formation of tissues and organs completely fails. As a consequence PKCi-deficient embryos die due to lack of a cardiovascular system. ESCs, when cultured as aggregates, form spherical structures which are defined as embryoid bodies. These structures are thought to recapitulate early steps of the pre-implantation development including endoderm formation, basement membrane assembly, epiblast polarization and subsequent cavity formation. When cultured in suspension for 5 days, EBs form epithelial cysts consisting of an outer endoderm and an inner columnar epiblast epithelium, separated by a BM. As PKCi mutants displayed an abnormal amniotic cavity at E7.5 we decided to use the embryoid body formation assay to analyze cavity formation in more detail. When wt and PKCi deficient ESCs were subjected to this assay, we detected obvious differences among the two genotypes. Wt EBs displayed the expected appearance in a phase contrast representation, PKCiD/D EBs showed a disorganized structure. Most of the wt EBs formed a single large cavity enclosed by CEE after 5 days, whereas the mutant EBs did not form a single cavity at all or formed multiple small cavities adjacent to the polarized CEE, although the endoderm differentiated normally. When cultured for additional 2 days, most mutant EBs formed small cavities but failed to complete cavitation. Since caspase-dependent NSC 136476 structure apoptosis has been described as a key mechanism involved in the early steps of cavity formation we immunostained 5-day EBs for cleaved caspase-3. Wt EBs displayed massive central apoptosis as evidenced by cleaved caspase-3 staining. By contrast, only scattered WY 14643 50892-23-4 apoptotic cells were detected in PKCi deficient EBs. DAPI staining of condensed and fragmented nuclei, another hallmark of apoptosis, also showed reduced apoptosis at the center of mutant EBs. To further analyze the apoptosis during EB cavitation, we cultured EBs of both genotypes for 2 to 5 days and performed immunoblotting for cleaved caspase-3. Indeed, PKCi deficient EBs expressed less apoptotic activity at all time points tested than the corresponding wt extracts. These results demonstrate that PKCi deficient EBs fail to form a single cavity possibly due to reduced central apoptosis. Gene targeting experiments have shown that laminin-mediated BM formation is essential for primitive ectoderm epithelialization and cavitation in EBs. The laminin a-1 chain is a key component of the embryonic BM expressed early on during development. In the wt EB the laminin a-1 staining formed a continuous thin layer between endoderm and epiblast cells. We did not observe obvious changes in BM assembly in mutant EBs. These findings were confirmed by using perlecan as an alternative marker for the BM. However, ectopic BM formation was detected at the center of,20% mutant EBs.
but subsequent formation of tissues and organs completely fails. As a consequence PKCi-deficient embryos die due to lack of a cardiovascular system. ESCs, when cultured as aggregates, form spherical structures which are defined as embryoid bodies. These structures are thought to recapitulate early steps of the pre-implantation development including endoderm formation, basement membrane assembly, epiblast polarization and subsequent cavity formation. When cultured in suspension for 5 days, EBs form epithelial cysts consisting of an outer endoderm and an inner columnar epiblast epithelium, separated by a BM. As PKCi mutants displayed an abnormal amniotic cavity at E7.5 we decided to use the embryoid body formation assay to analyze cavity formation in more detail. When wt and PKCi deficient ESCs were subjected to this assay, we detected obvious differences among the two genotypes. Wt EBs displayed the expected appearance in a phase contrast representation, PKCiD/D EBs showed a disorganized structure. Most of the wt EBs formed a single large cavity enclosed by CEE after 5 days, whereas the mutant EBs did not form a single cavity at all or formed multiple small cavities adjacent to the polarized CEE, although the endoderm differentiated normally. When cultured for additional 2 days, most mutant EBs formed small cavities but failed to complete cavitation. Since caspase-dependent NSC 136476 structure apoptosis has been described as a key mechanism involved in the early steps of cavity formation we immunostained 5-day EBs for cleaved caspase-3. Wt EBs displayed massive central apoptosis as evidenced by cleaved caspase-3 staining. By contrast, only scattered WY 14643 50892-23-4 apoptotic cells were detected in PKCi deficient EBs. DAPI staining of condensed and fragmented nuclei, another hallmark of apoptosis, also showed reduced apoptosis at the center of mutant EBs. To further analyze the apoptosis during EB cavitation, we cultured EBs of both genotypes for 2 to 5 days and performed immunoblotting for cleaved caspase-3. Indeed, PKCi deficient EBs expressed less apoptotic activity at all time points tested than the corresponding wt extracts. These results demonstrate that PKCi deficient EBs fail to form a single cavity possibly due to reduced central apoptosis. Gene targeting experiments have shown that laminin-mediated BM formation is essential for primitive ectoderm epithelialization and cavitation in EBs. The laminin a-1 chain is a key component of the embryonic BM expressed early on during development. In the wt EB the laminin a-1 staining formed a continuous thin layer between endoderm and epiblast cells. We did not observe obvious changes in BM assembly in mutant EBs. These findings were confirmed by using perlecan as an alternative marker for the BM. However, ectopic BM formation was detected at the center of,20% mutant EBs.
Despite differences in the stages that sequester in both species adhere to the vascular endothelium via CD36
If no SUMOylation is found, then a heterologous co-expression of SUMO-1 and SERCA2 should be considered. With regard to the solubilisation detergent, as tested in the case of the hSERCA2a-GFP-His8 construct, no increase in the yield of purified protein was observed when using a smaller detergent: total membrane protein ratio. Thus, it can be concluded that the initial 3:1 ratio used did not contribute to the high aggregation observed during SEC. Further, based on the result obtained with FC12, it may be that usage of a higher solubility capacity detergent may lead to aggregated fraction solubilisation rather than the solubilisation of the active form of the protein. Further work would be necessary to more fully understand the effect of different detergents or lipids on human cardiac Ca2+-ATPase stability. Purified hSERCA2a showed calcium-dependent and thapsigargin-sensitive activity. The calcium K0.5 for hSERCA2a of 0.6 ��M found herein is within previously reported values. A significant difference observed in the turn-over rate between previously purified samples of SERCA2a may be explained by the different lipid content, since previous data were measured on vesicular microsomes which contain some natural lipids associated with SERCA2, whereas SERCA2a purified from S. cerevisiae was not reconstituted into liposomes and was analysed in the presence of detergent. Another plausible explanation for the differences in turn-over rate could be the presence of inactive protein in the final purified sample. Finally and importantly, are the diversity of ATPase assay conditions, which may explain the significant differences in the enzymatic activities reported. It is noteworthy that, although the turn-over rate for hSERCA2a was different than when expressed using other systems, the values obtained here are very close to the specific enzymatic activities obtained for rSERCA1a expressed and purified from S. cerevisiae. The optimised protocol Niltubacin side effects outlined in this work is easily extended to other SERCA isoforms and useful for the production of high quality recombinant active protein for further analysis to study interactions between SERCAs and their physiologically relevant partners. The resulting protein is suitable for crystallisation trials and subsequent structural analysis. Furthermore, the method outlined may prove useful generally for the recombinant production of other multi-domain eukaryotic membrane proteins. The etiological agents of human malaria are vector-borne protozoan parasites that initially infect liver cells but rapidly develop to invade and reproduce in host erythrocytes. These blood-stage parasites produce a number of proteins that are exported to the surface of the infected red blood cell. The pattern of adherence is selective in that most sequestered infected iRBCs are found in the lungs, adipose tissue, brain and placenta. The ability to sequester in the vasculature of certain tissues is thought to be advantageous to the parasite because it diminishes clearance of trophozoite- and schizont-containing iRBCs in the spleen and promotes factors that are beneficial for parasite growth. Sequestration in organs such as the lung is not without consequences, as large numbers of iRBCs in the lung have been hypothesized to precipitate events 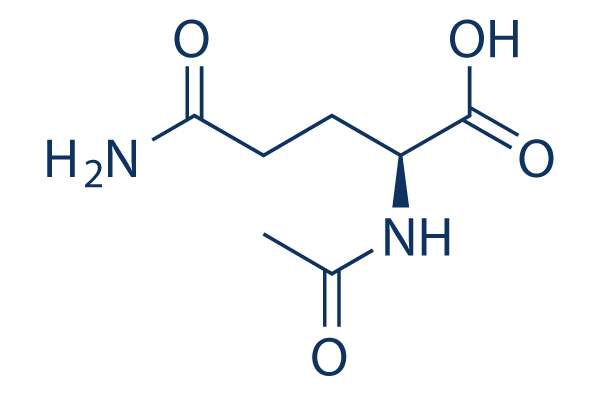 that result in lung injury. Work with synchronized PbA infections has demonstrated that schizontcontaining RBCs selectively adhere to lung microvascular endothelial cells and this interaction is dependent on capillary endothelial cell expression of CD36. Interestingly, the PbA genome does not contain any PfEMP1 orthologues. Recently, it has been demonstrated that the schizont membraneassociated cytoadherence protein is critical for PbA schizont-stage parasites to adhere to vascular Torin 1 endothelium via CD36.
that result in lung injury. Work with synchronized PbA infections has demonstrated that schizontcontaining RBCs selectively adhere to lung microvascular endothelial cells and this interaction is dependent on capillary endothelial cell expression of CD36. Interestingly, the PbA genome does not contain any PfEMP1 orthologues. Recently, it has been demonstrated that the schizont membraneassociated cytoadherence protein is critical for PbA schizont-stage parasites to adhere to vascular Torin 1 endothelium via CD36.
We tested the reliability of our TIS prediction for the alternative proteins previously detecte
A total of 68,035 peptides from 5,558 reference proteins and 280 peptides from 129 alternative proteins were identified. The mean sequence coverage for reference and alternative proteins was 28.8% and 32.3%, respectively. Overall, alternative proteins represented 2.27% of the total identified proteins. This result clearly shows that the contribution of alternative proteins to the proteome, and thus the number of multiple coding genes, has been overlooked. It is noteworthy that alternative proteins coding sequences are spread across the different regions of mRNAs in agreement with the predicted distribution. Co-Tulathromycin B expression of an alternative protein and its reference protein was observed for 42 genes. For each of these genes, the average peptide intensity plot of both the reference and alternative proteins revealed large variations in co-expression ratio, indicating that a reference protein might not always be the main protein product of a gene. To confirm the expression of alternative proteins in cell lines different from HeLa cells, we performed LCMS/MS on human colon cell lines and identified 45 alternative proteins. AltORFs associated with these 45 proteins were distributed within UTRs and RefORFs with frequencies comparable to those observed in HeLa cells. Comparative analysis of alternative proteins detected in both HeLa cells and colon cell lines indicated that 14 are expressed in at least two cell lineages. This is more than expected by chance. SDS-PAGE in combination with LC-MS/MS is generally limited to the analysis of proteins above 10 kDa, and a low molecular weight is a known limitation in protein identification by MS. Since the majority of the predicted alternative proteome is composed of proteins less than 90 amino acids long which have a predicted molecular weight below 10 kDa, it is not surprising to have detected much more peptides corresponding to the conventional proteome compared to the alternative proteome. To further assess the abundance of the alternative proteome compared to the conventional proteome, HeLa cells proteins were separated by 1-D SDS-PAGE, and one gel slice between the 4.6 and 10 kDa markers was trypsin digested. The resulting peptides were analyzed by LC-MS/MS. A total of 44 reference and 14 alternative proteins were detected, and alternative proteins represented 24.14% of the total identified proteins, thus showing that alternative proteins are enriched in the pool of small cellular proteins. The detection of alternative proteins with MW between 4.78 and 9.49 kDa is further proof that peptides were not misassigned and that these alternative proteins are actually expressed. Next, we tested the expression of alternative proteins in a variety of human tissues by LC-MS/MS. First, we analyzed normal colon and lung tissues and detected 13 and 40 alternative proteins respectively. In a second set of experiments, we analyzed ovarian cancer tissue areas and normal areas from the same formalin fixed, paraffin-embedded tissue section of two patients, one presenting endometrioid ovarian cancer and the second presenting a serous ovarian cancer. A total of 19 alternative proteins were identified in the normal endometrium, endometrioid ovary, serous ovary, normal ovary, and serous fallopian tube. We completed these proteomic studies with human fluids, including cerebrospinal fluid, urine, plasma, and serum, identifying 16, 47, 90, and 928 alternative proteins in each fluid respectively. Strikingly, alternative proteins reButenafine hydrochloride present approximately 55% of the proteins identified in plasma and serum. Overall, we detected a total of 1,259 alternative proteins, and 47 were expressed in different cell lines and/or tissues. In accordance with the scanning model of translation initiation, we used the first AUG rule in order to predict ![]() the TIS of AltORFs present in our database.
the TIS of AltORFs present in our database.
It is becoming increasingly apparent that nuclear HER family transcriptional functions in cancer biology
These functions may be Mepiroxol independent of their 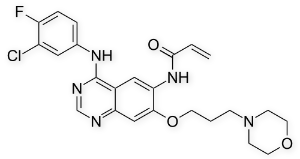 kinase activity. The current study supports these findings because 1) HER3 lacks high levels of kinase activity, and 2) nuclear HER3 regulation of cyclin D1 was independent of plasma membrane-bound HER3 functions and AKT signaling. Since the kinase functions of HER family members may not be solely responsible for the tumorigenic properties of these receptors, the identification of HER family TADs may serve as a map to better target nuclear HER functions in the future. Four members have ATP-independent chaperone activity but only three of them display enhanced level of expression in 3,4,5-Trimethoxyphenylacetic acid response to heat shock or stimuli that misfold and damage polypeptides. sHsps are constitutively expressed in many different tissues. In that regard, HspB1 is expressed in most tissues. HspB5 is also expressed in a wide range of tissues, including lens, heart, skeletal muscle, colon, lung and kidney whereas HspB4 is mainly expressed in the lens. These proteins have the ability to interact with each other and form hetero-oligomeric complexes. For example, in mammalian lenses, HspB4 and HspB5 form a major structural protein complex, denoted a-crystallin, involved in the refractive and light focusing properties of the lens. This complex is present in both the water soluble and insoluble fractions of the lens, especially in the nucleus region, a domain of the lens where the only other detectable sHsp, particularly in caracteous lens, is the phosphorylated form of HspB1. In addition, these Hsps share the ability to enhance the resistance of cells to the deleterious effects induced by stresses, such as those induced by heat shock, drugs, UV light and alterations in intracellular redox homeostasis. In that respect, they prevent aggregation and precipitation of misfolded or oxidized proteins. In addition, HspB1 and HspB5 can act as antioxidant proteins leading to the establishment of a pro-reducing state in cells by up-regulating the activity of antioxidant enzymes, such as glucose 6-phosphate dehydrogenase. These two chaperones also play anti-apoptotic and tumorigenic roles by interacting with specific key protein partners and are nowadays considered as potent anti-cancer therapeutic targets. Another major role of HspB1, HspB4 and HspB5 relates to their ability to modulate and stabilize cytoskeleton architecture. For instance, HspB5 chaperone activity is required to stabilize and modulate intermediate filaments assembly and avoid their aggregation. In that respect, several mutations in HspB5 have been shown to alter cytoskeletal architecture, such as the natural missense mutation R120G, which is responsible for cataracts, cardiomyopathies and desmin-related myopathies. The removal of the positive charge from arginine 120 is known to cause HspB5 partial unfolding, increased exposure of hydrophobic regions, abnormal assemblies and subunit exchange and enhanced susceptibility to proteolysis. The mutation also reduces HspB5 solubility and promotes its aggregation. In addition, it strongly impairs HspB5 chaperone activity. A fundamental property of sHsps is their ability to oligomerize. For example, HspB1 forms dynamic polydispersed structures with heterogenous native sizes comprised between 50 and 800 kDa while HspB5 native size is more uniformly distributed within the 700 to 800 kDa range. The oligomerization of HspB1 is a dynamic phenomenon linked to cell physiology that probably allows HspB1 interaction with specific client proteins. On the other hand, HspB1 large oligomers can also act as reservoirs that store stress-induced misfolded or oxidized polypeptides until they are either refolded by ATP-dependent chaperones or degraded. Another parameter to take into account is the phosphorylation of sHsps. This is a complex phenomenon because these proteins have several serine sites that can be phosphorylated different.
kinase activity. The current study supports these findings because 1) HER3 lacks high levels of kinase activity, and 2) nuclear HER3 regulation of cyclin D1 was independent of plasma membrane-bound HER3 functions and AKT signaling. Since the kinase functions of HER family members may not be solely responsible for the tumorigenic properties of these receptors, the identification of HER family TADs may serve as a map to better target nuclear HER functions in the future. Four members have ATP-independent chaperone activity but only three of them display enhanced level of expression in 3,4,5-Trimethoxyphenylacetic acid response to heat shock or stimuli that misfold and damage polypeptides. sHsps are constitutively expressed in many different tissues. In that regard, HspB1 is expressed in most tissues. HspB5 is also expressed in a wide range of tissues, including lens, heart, skeletal muscle, colon, lung and kidney whereas HspB4 is mainly expressed in the lens. These proteins have the ability to interact with each other and form hetero-oligomeric complexes. For example, in mammalian lenses, HspB4 and HspB5 form a major structural protein complex, denoted a-crystallin, involved in the refractive and light focusing properties of the lens. This complex is present in both the water soluble and insoluble fractions of the lens, especially in the nucleus region, a domain of the lens where the only other detectable sHsp, particularly in caracteous lens, is the phosphorylated form of HspB1. In addition, these Hsps share the ability to enhance the resistance of cells to the deleterious effects induced by stresses, such as those induced by heat shock, drugs, UV light and alterations in intracellular redox homeostasis. In that respect, they prevent aggregation and precipitation of misfolded or oxidized proteins. In addition, HspB1 and HspB5 can act as antioxidant proteins leading to the establishment of a pro-reducing state in cells by up-regulating the activity of antioxidant enzymes, such as glucose 6-phosphate dehydrogenase. These two chaperones also play anti-apoptotic and tumorigenic roles by interacting with specific key protein partners and are nowadays considered as potent anti-cancer therapeutic targets. Another major role of HspB1, HspB4 and HspB5 relates to their ability to modulate and stabilize cytoskeleton architecture. For instance, HspB5 chaperone activity is required to stabilize and modulate intermediate filaments assembly and avoid their aggregation. In that respect, several mutations in HspB5 have been shown to alter cytoskeletal architecture, such as the natural missense mutation R120G, which is responsible for cataracts, cardiomyopathies and desmin-related myopathies. The removal of the positive charge from arginine 120 is known to cause HspB5 partial unfolding, increased exposure of hydrophobic regions, abnormal assemblies and subunit exchange and enhanced susceptibility to proteolysis. The mutation also reduces HspB5 solubility and promotes its aggregation. In addition, it strongly impairs HspB5 chaperone activity. A fundamental property of sHsps is their ability to oligomerize. For example, HspB1 forms dynamic polydispersed structures with heterogenous native sizes comprised between 50 and 800 kDa while HspB5 native size is more uniformly distributed within the 700 to 800 kDa range. The oligomerization of HspB1 is a dynamic phenomenon linked to cell physiology that probably allows HspB1 interaction with specific client proteins. On the other hand, HspB1 large oligomers can also act as reservoirs that store stress-induced misfolded or oxidized polypeptides until they are either refolded by ATP-dependent chaperones or degraded. Another parameter to take into account is the phosphorylation of sHsps. This is a complex phenomenon because these proteins have several serine sites that can be phosphorylated different.